
El fascinante viaje de las proteínas: ¿Cómo navegan por la célula eucariota?
Un recorrido por las sofisticadas rutas y mecanismos que transportan estas moléculas esenciales a sus destinos celulares
Aspectos fundamentales del transporte proteico
- Sistema de direccionamiento preciso: Las proteínas contienen "códigos postales" moleculares que determinan su destino final dentro o fuera de la célula
- Red logística integrada: El transporte involucra múltiples orgánulos coordinados que procesan, modifican y dirigen las proteínas
- Mecanismos especializados: Diferentes vías de transporte aseguran que cada proteína llegue exactamente donde se necesita para su función específica
El transporte de proteínas en células eucariotas es un proceso extraordinariamente sofisticado y esencial para la vida celular. A diferencia de las células procariotas más simples, las eucariotas poseen compartimentos internos especializados que requieren un tráfico proteico preciso y regulado. Veamos cómo estas moléculas esenciales viajan desde su síntesis hasta su destino final.
Inicio del viaje: Síntesis proteica y entrada al sistema de transporte
El viaje de las proteínas comienza con su síntesis en los ribosomas, las "fábricas proteicas" celulares. La ubicación de esta síntesis determina en gran medida la ruta que seguirán posteriormente:
Ribosomas libres vs. ribosomas asociados al retículo endoplasmático
Existen dos poblaciones principales de ribosomas en la célula eucariota:
- Ribosomas libres en el citosol: Sintetizan proteínas destinadas a permanecer en el citoplasma, núcleo, mitocondrias, cloroplastos y peroxisomas
- Ribosomas asociados al retículo endoplasmático rugoso (RER): Producen proteínas que serán secretadas al exterior celular o incorporadas a membranas celulares
El direccionamiento al RER ocurre gracias a una secuencia señal en el extremo N-terminal de la proteína naciente. Esta secuencia es reconocida por una partícula de reconocimiento de señal (SRP) que guía al complejo ribosoma-proteína hacia el RER, donde ocurre la translocación cotraduccional a través del complejo proteico Sec61.
Translocación y procesamiento en el RER
Una vez en el RER, las proteínas experimentan su primera fase de procesamiento:
- Plegamiento asistido por chaperonas para adquirir su estructura tridimensional correcta
- Formación de enlaces disulfuro para estabilizar la estructura proteica
- Glicosilación inicial mediante la adición de oligosacáridos a residuos de asparagina (N-glicosilación)
- Control de calidad que asegura el correcto plegamiento antes de avanzar en la vía secretora
El gráfico anterior muestra una comparación entre diferentes vías de transporte proteico, evaluando factores como complejidad, especificidad y gasto energético. La vía secretora (RE-Golgi) destaca por su alta especificidad y diversidad de proteínas transportadas, mientras que otras rutas como el transporte a mitocondrias o núcleo tienen sus propias características distintivas.
La vía secretora: Tránsito por el aparato de Golgi
El aparato de Golgi representa la "central de clasificación" de las proteínas en la célula eucariota. Las proteínas que salen del RE son transportadas al Golgi mediante vesículas recubiertas de proteínas COPII.
Estructura y función del aparato de Golgi
El aparato de Golgi está organizado en cisternas apiladas que definen tres regiones principales:
- Cara cis: Recibe vesículas desde el RE y representa la entrada al Golgi
- Región medial: Zona intermedia donde ocurren numerosas modificaciones
- Cara trans: Región de salida donde las proteínas son clasificadas y empaquetadas en vesículas según su destino final
Procesamiento proteico en el Golgi
Durante su tránsito por el Golgi, las proteínas experimentan diversas modificaciones:
- Modificación de los oligosacáridos añadidos en el RE
- Adición de nuevos azúcares (O-glicosilación)
- Sulfatación de determinados residuos
- Fosforilación de aminoácidos específicos
- Procesamiento proteolítico (corte de segmentos específicos)
El tráfico vesicular: Mecanismo fundamental del transporte proteico
El transporte entre compartimentos celulares se realiza principalmente mediante vesículas. Este proceso incluye varias etapas clave:
- Formación de vesículas: Reclutamiento de proteínas de cubierta (COPI, COPII o clatrina) que deforman la membrana
- Captura de carga: Inclusión selectiva de proteínas específicas en la vesícula naciente
- Gemación y escisión: Separación de la vesícula de la membrana donante
- Transporte: Movimiento de la vesícula a través del citoplasma, facilitado por el citoesqueleto
- Anclaje: Reconocimiento específico de la membrana diana mediante proteínas de anclaje
- Fusión: Incorporación de la vesícula a la membrana diana mediante proteínas SNARE
Este video explica detalladamente el proceso de tráfico de proteínas entre los diferentes compartimentos celulares, mostrando cómo las vesículas transportan proteínas entre orgánulos y hacia el exterior celular.
Destinos finales: Clasificación y direccionamiento
Después de procesarse en el Golgi, las proteínas son clasificadas y enviadas a diferentes destinos. Este proceso es mediado por señales específicas en las proteínas y receptores especializados.
| Destino | Mecanismo de transporte | Señal de direccionamiento | Ejemplo de proteínas |
|---|---|---|---|
| Membrana plasmática | Vesículas de secreción constitutiva | Señales de transmembrana, modificaciones lipídicas | Receptores, transportadores, proteínas de adhesión |
| Secreción al exterior | Vesículas de secreción regulada o constitutiva | Péptido señal, ausencia de señales de retención | Hormonas, enzimas digestivas, anticuerpos |
| Lisosomas | Vesículas recubiertas de clatrina | Manosa-6-fosfato (M6P) | Hidrolasas ácidas, proteasas lisosomales |
| Endosomas | Vesículas recubiertas de clatrina | Señales basadas en tirosina o dileucina | Receptores de reciclaje, proteínas de clasificación |
| Mitocondrias | Translocación postraduccional | Secuencia de localización mitocondrial (MLS) | Proteínas de la cadena respiratoria, enzimas metabólicas |
| Núcleo | Transporte a través del complejo del poro nuclear | Secuencia de localización nuclear (NLS) | Factores de transcripción, histonas, polimerasas |
| Peroxisomas | Translocación postraduccional | Secuencia de localización peroxisomal (PTS1, PTS2) | Catalasa, enzimas oxidativas |
Transporte a orgánulos no vinculados a la vía secretora
Algunos orgánulos, como mitocondrias, cloroplastos y núcleo, reciben proteínas mediante mecanismos diferentes a la vía secretora clásica:
Transporte a mitocondrias y cloroplastos
Las proteínas destinadas a estos orgánulos se sintetizan en ribosomas libres y contienen secuencias de localización específicas. Son reconocidas por complejos de translocación especializados en la membrana externa de estos orgánulos (complejos TOM/TIM para mitocondrias y TOC/TIC para cloroplastos), que facilitan su importación.
Transporte al núcleo
El transporte nuclear ocurre a través de los complejos del poro nuclear (NPC). Las proteínas con secuencias de localización nuclear (NLS) son reconocidas por importinas que las transportan al interior del núcleo mediante un proceso dependiente de la GTPasa Ran.
El papel del citoesqueleto en el transporte de proteínas
El citoesqueleto proporciona las "vías" por las que se mueven las vesículas y orgánulos. Existen dos sistemas principales:
- Microtúbulos: Sirven como "carreteras" de larga distancia. El transporte ocurre mediante proteínas motoras (quinesinas y dineínas) que se mueven en direcciones específicas
- Filamentos de actina: Facilitan el transporte a corta distancia, especialmente cerca de la membrana plasmática, mediante proteínas motoras como las miosinas
Visualización del transporte proteico en células eucariotas
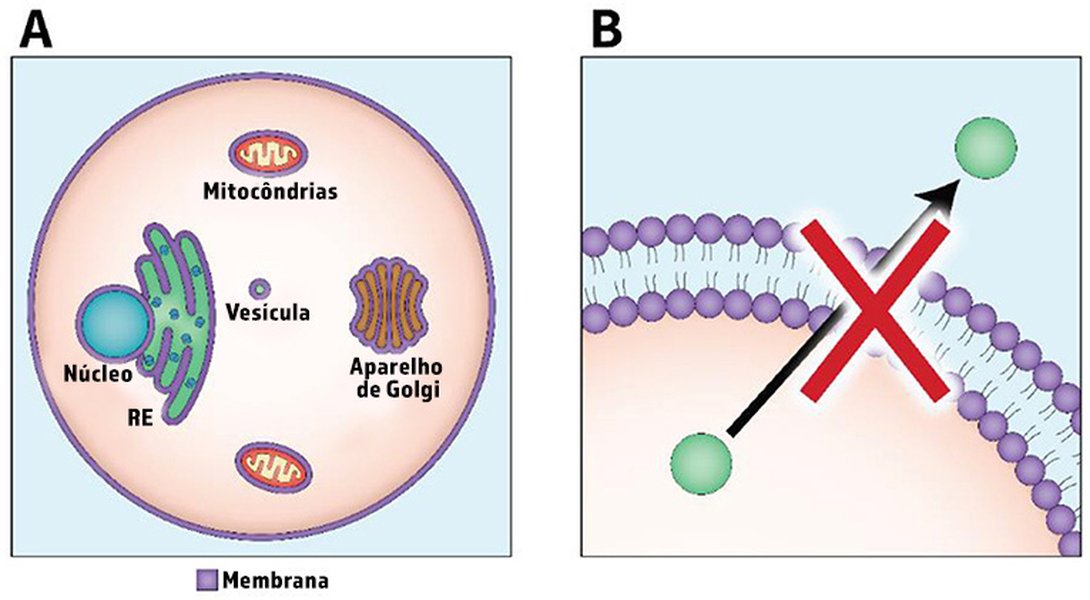
Representación del sistema de transporte proteico en una célula eucariota, mostrando el retículo endoplasmático, aparato de Golgi y vesículas de transporte.

Detalle del proceso de secreción proteica, desde la síntesis en el RER hasta la liberación al exterior celular.
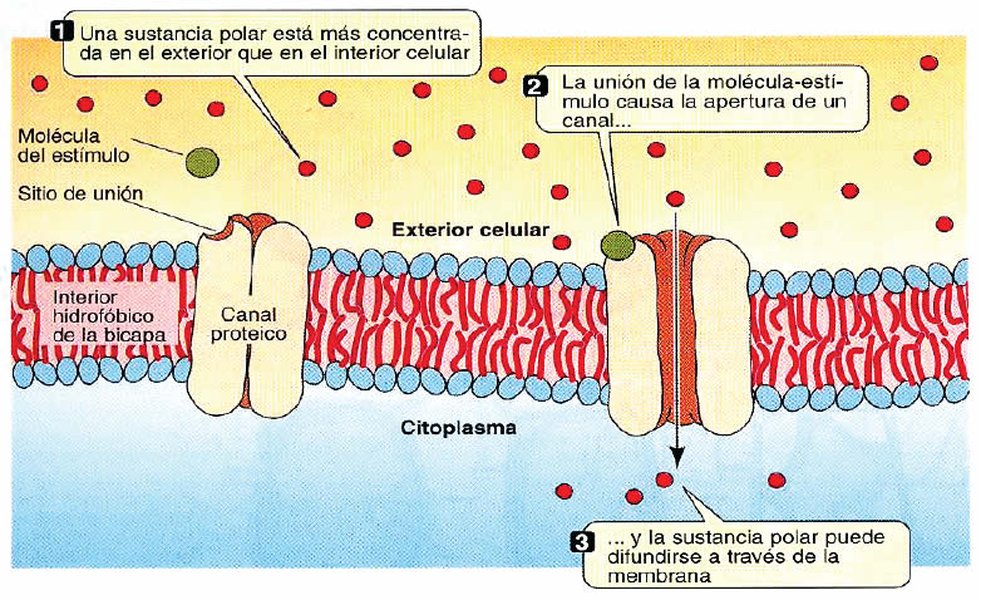
Ilustración que muestra el transporte facilitado por proteínas a través de la membrana celular.
Mapa mental: Rutas de transporte proteico en células eucariotas
A continuación se presenta un mapa mental que resume las principales vías de transporte de proteínas en la célula eucariota, desde su síntesis hasta su destino final:
en células eucariotas"] ["Síntesis proteica"] ["Ribosomas libres"] ["Proteínas para citosol"] ["Proteínas para núcleo"] ["Proteínas para mitocondrias"] ["Proteínas para peroxisomas"] ["Ribosomas asociados al RE"] ["Proteínas de secreción"] ["Proteínas de membrana"] ["Proteínas lisosomales"] ["Vía secretora"] ["Retículo endoplasmático"] ["Plegamiento"] ["Glicosilación inicial"] ["Control de calidad"] ["Aparato de Golgi"] ["Modificaciones adicionales"] ["Clasificación y empaquetamiento"] ["Destinos postGolgi"] ["Membrana plasmática"] ["Secreción"] ["Endosomas/Lisosomas"] ["Transporte postraduccional"] ["Mitocondrias (TOM/TIM)"] ["Cloroplastos (TOC/TIC)"] ["Núcleo (NPC)"] ["Peroxisomas (PEX)"] ["Mecanismos de transporte"] ["Transporte vesicular"] ["COPII (RE→Golgi)"] ["COPI (Golgi→RE)"] ["Clatrina (Golgi→Lisosomas)"] ["Translocación a través de membranas"] ["Transporte mediado por receptores"]
Preguntas frecuentes sobre el transporte de proteínas
Referencias
- Ribosomas: estructura, función y papel en la traducción - Genotipia
- ¿Qué es la síntesis de proteínas? - Cromtek
- Transporte de proteínas: Mecanismos, rutas y regulación celular - Kobu
- Transporte de proteínas desde el retículo endoplasmático al aparato de Golgi - Universidad de Vigo
- La célula: Tráfico vesicular - Atlas de Histología Vegetal y Animal
- Proteínas de Transporte: definición, tipos, función y ejemplos - Estudyando
- Transporte celular: tipos y sus características - Lifeder
- Transporte de proteínas a la mitocondria y cloroplasto - Wikibooks
Lecturas recomendadas
 cajalesygalileos.wordpress.com
cajalesygalileos.wordpress.com
Last updated April 7, 2025